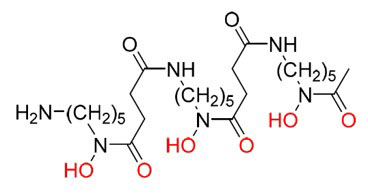
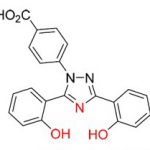
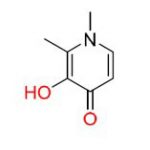
[1] Une macromolécule est une molécule possédant une masse molaire élevée. Dans l’organisme vivant, il s’agit de polymères, c’est-à-dire des molécules formées par l’enchaînement d’une unité de répétition appelée monomère : protéines (enchaînements d’acides aminés), ADN (enchaînement d’acides nucléiques), ou encore polysaccharides (enchaînements de monosaccharides tels que le glucose) comme l’amidon ou la cellulose.
[2] Les protéines sont des macromolécules réalisant des fonctions biologiques diverses dans l’organisme : protéines structurelles, protéines de transport (par exemple les protéines transmembranaires, qui transportent des molécules entre le milieu intracellulaire et le milieu extracellulaire), protéines de régulation, protéines de signalisation (hormones…), protéines réceptrices, sensorielles, protéines motrices, protéines de défense (anticorps…), protéines de stockage et enzymes.
[3] Un cofacteur est une molécule qui est nécessaire à l’activité biologique d’une protéine, en général une enzyme. Il peut s’agir d’un ion métallique ou d’une molécule telle que les vitamines.
[4] Cytotoxique : toxique pour les cellules.
 Présentation
Présentation  Entités
Entités  Partenaires
Partenaires  Histoire de la SCF
Histoire de la SCF  Gouvernance
Gouvernance  Industrie
Industrie 
{kind=link}
{kind=link}
{kind=link}
{kind=link}


{kind=link}